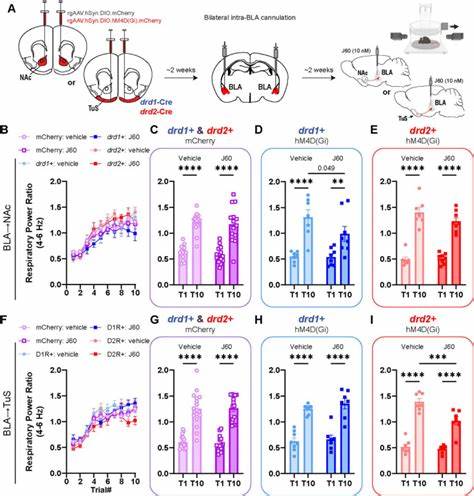
Негативные эмоциональные состояния, такие как страх, тревога и избегание, составляют фундаментальные реакции организма на потенциальную угрозу и играют ключевую роль в адаптивном поведении. Однако дисфункции в процессах их формирования или регуляции могут привести к развитию серьезных психических расстройств, включая тревожные расстройства и посттравматическое стрессовое расстройство. Центральным элементом нервной системы, отвечающим за обработку эмоциональных стимулов, является базолатеральный комплекс миндалины (БЛМ), который служит интеграционным центром для восприятия и интерпретации сенсорной информации с последующей генерацией поведенческих и эмоциональных реакций. Современные исследования показывают, что БЛМ состоит из гетерогенных нейронных популяций, различающихся по генетической маркерной структуре и функциональным связям с другими областями мозга, что открывает путь к пониманию того, как именно формируются различные эмоциональные состояния. Одним из важнейших обновлений в понимании функционирования БЛМ стало обнаружение двух параллельных генетически различных путей, идущих от нейронов, экспрессирующих рецепторы дофамина D1 (Drd1+) и D2 (Drd2+), к различным подрегиональным структурам вентрального стриатума, таким как ядро Accumbens (NAc) и трубчатое ядро (TuS).
Эти параллельные пути не только анатомически разделены, но и функционально направлены на регуляцию различных аспектов эмоционального поведения, включая как врожденные, так и приобретённые реактивные состояния. Роль этих двух путей была детально изучена в серии поведенческих экспериментов с применением современных методов, таких как оптогенетика и хемогенетика, что позволило понять вклад каждого пути в модуляцию негативных эмоций. Базолатеральный комплекс миндалины содержит различные типы клеток, среди которых особый интерес вызывают нейроны, экспрессирующие Drd1 и Drd2 – гены, кодирующие рецепторы дофамина D1 и D2 соответственно. Эти рецепторы участвуют в регуляции внутриклеточных сигналов через путь фосфорилирования и дефосфорилирования белка PPP1R1B (также известного как DARPP-32), который интегрирует допаминергические сигналы. Почти все дофаминергические сигналы в БЛМ направлены на эти два основных класса нейронов, и они формируют два параллельных, но функционально различимых проекционных пути к вентральному стриатуму.
Первый путь состоит из Drd1+ нейронов, которые преимущественно проецируются на ядро Accumbens. Активация этой цепи ассоциируется с увеличением избегания и усилением негативной эмоциональной реакции на определенные стимулы. Оптогенетическое стимулирование терминалей Drd1+ нейронов в ядре Accumbens приводит к появлению поведенческих признаков избегания в реальном времени, что свидетельствует о том, что данный путь способен быстро и эффективно модулировать эмоциональное состояние животного. Второй путь включает Drd2+ нейроны БЛМ, которые преимущественно проецируются на трубчатое ядро вентрального стриатума. В отличие от первого пути, стимуляция Drd2+ нейронов, направленных на TuS, также вызывает поведенческое избегание, но в другом контексте и с иной динамикой регуляции приобретаемых эмоциональных реакций.
Эти две проекционные системы, хотя и формируют параллельные пути с общими целями – регулировать негативные эмоции, – имеют уникальные и интригующие различия, которые проявляются в способах их воздействия на поведение и кодировании эмоционального восприятия. Кроме того, исследования с использованием хемогенетической инактивации позволили продемонстрировать важность каждого из этих путей в формировании условных ассоциаций страха в павловской модели. При подавлении активности Drd1+ проекций в ядро Accumbens и Drd2+ проекций в TuS наблюдалось снижение интенсивности выученного страха, выраженного как в поведении (подвижность, замерзание), так и в физиологических реакциях, включая параметры дыхания, что подчеркивает их ключевую роль в модуляции негативных эмоциональных состояний на уровне обучения. Другим интересным открытием является то, что в вентральном стриатуме, как и в БЛМ, существуют две основные категории спинальных проекционных нейронов – Drd1+ и Drd2+, которые получают возбуждающие синаптические входы от соответствующих Drd1+ и Drd2+ БЛМ нейронов. Эксперименты in vitro показали, что активация терминалей Drd1+ БЛМ нейронов вызывает более мощные моносинаптические возбуждающие постсинаптические потенциалы в Drd1+ нейронах TuS по сравнению с их воздействием на Drd2+ нейроны.
Наоборот, Drd2+ БЛМ нейроны активируют Drd1+ и Drd2+ нейроны TuS в большей степени в ненаправленном режиме и с меньшей амплитудой возбуждения. Такая функциональная организация может обеспечивать селективное влияние на выходные цепи вентрального стриатума, играющего центральную роль в мотивации, награде и эмоциональной регуляции. Уникальность и параллельность путей Drd1+ и Drd2+ нейронов БЛМ создают сложную нейронную сеть, способную обеспечить гибкое и контекстуально-зависимое регулирование эмоциональных и поведенческих реакций. Такие данные расширяют традиционные представления о роли амидалины исключительно как центра страха и подтверждают, что БЛМ оказывает многоаспектное влияние на эмоциональную мотивацию через прямые и проекционно-специфичные взаимодействия с вентральным стриатумом. Учитывая обнаруженную гетерогенность данных проекций, можно предположить, что нарушение баланса активности между этими двумя путями может лежать в основе патогенеза тревожных и стрессовых расстройств.
В частности, гиперактивность Drd1+ проекций в NAc или Drd2+ проекций в TuS может приводить к усиленному формированию и выражению негативных эмоций. В связи с этим нацеливание на эти специфичные нейронные пути и их рецепторное окружение может стать перспективным направлением для разработки новых терапевтических методов с более высокой избирательностью и эффективностью, чем современные подходы. Подытоживая, современные исследования нейродинамики эмоциональных состояний выявили параллельные пути из базолатерального миндалина в вентральный стриатум, отражающие генетическую и функциональную специфику нейрональных субпопуляций, влияющих на регуляцию негативных эмоций. Эти данные усиливают наше понимание сложной организации эмоциональных цепей мозга и предлагают новые перспективы для изучения механизмов тревожных и связанных расстройств. Последующие исследования будут необходимы для детального понимания способов взаимодействия этих путей с другими нейронными сетями, такими как центральный миндалевидный комплекс, вентральное тегментальное ядро и префронтальная кора.
Также важным направлением является изучение того, как нейромодуляторы, особенно дофамин, регулируют эти пути в различных эмоциональных контекстах, что позволит раскрыть динамические механизмы эмоциональной пластичности и адаптации на молекулярном и системном уровне. В целом, глубокое понимание работы параллельных генетически определенных проекционных путей из базолатерального миндалина к подрегиональным структурам вентрального стриатума обогатит нейронаучные подходы к диагностике и лечению эмоциональных расстройств, открывая двери к инновационным вмешательствам, основанным на точной модуляции нервных цепей.