Эволюция многоклеточности стала ключевым поворотным моментом в развитии жизни на Земле, открыв путь к появлению более крупных и сложных организмов. Увеличение размера зачастую приносит многочисленные преимущества — защиту от хищников, повышение эффективности использования ресурсов и возможность освоения новых экологических ниш. Однако с ростом размера возникают и серьезные биофизические проблемы, одна из самых значительных из которых связана с транспортом питательных веществ и удалением отходов. В частности, диффузия — пассивный процесс перемещения веществ — становится недостаточным для обеспечения жизнедеятельности клеток, расположенных в глубине многоклеточных скоплений. Именно поэтому многие современные крупные организмы развили специализированные системы доставки питательных веществ, такие как сосудистые сети или двигательные структуры, генерирующие активные потоки вокруг и внутри организма.
Но как могли преодолевать подобные ограничения первые многоклеточные формы жизни, не обладавшие такими сложными адаптациями? Современные исследования, особенно в области экспериментальной эволюционной биологии, дают ответы и неожиданные перспективы на этот вопрос. Одним из уникальных объектов исследования служат снежные дрожжи — модельная система, в которой простейшие одноклеточные дрожжи Saccharomyces cerevisiae экспериментально превращаются в многоклеточные колонии посредством естественного отбора и серии эволюционных экспериментов. В ходе длительной селекции эти колонии достигают размера в миллиметры — что для дрожжей цифра гигантская. В таких масштабах традиционные представления о транспортных процессах через диффузию перестают работать, и предполагалось, что рост подобных скоплений должен быть ограничен исключительно поверхностью, что ведёт к замедлению и субэкспоненциальному росту. Однако экспериментальные наблюдения удивили учёных — снежные дрожжевые кластеры продолжают расти экспоненциально, даже достигая макроскопических размеров.
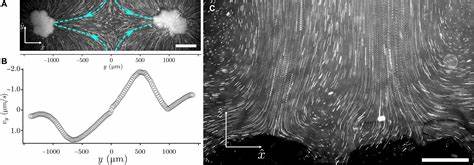
Главным открытием стал факт возникновения вокруг и внутри многоклеточных скоплений активных потоков жидкости, сопоставимых по скорости с потоками, создаваемыми специализированными органоидами в более сложных многоклеточных организмах, например, ресничками и жгутиками. Гидродинамические измерения показали циркуляционные движения жидкости, где ее движение входит с боков и выходит сверху кластеров. Изначально такой процесс ассоциировался с активным биологическим движением, однако снежные дрожжи не имеют специализированных структур вроде ресничек, порождающих эти потоки. Раскрылась более тонкая физическая картина: метаболическая активность колоний изменяет локальную плотность окружающей среды через насыщение продукта ферментации — этанола и углекислого газа — которые менее плотные, чем исходная среда, вызывая градиенты плотности. Эти градиенты, под воздействием силы тяжести, индуцируют естественные конвективные потоки («тепловые» или «плотностные»), способствующие более эффективному перемещению питательных веществ к внутренним клеткам и распределению продуктов обмена.
Подтверждение механизма пришло из экспериментов, в которых использовались флуоресцентные микроскопические частицы для визуализации потоков. Учёные зафиксировали различие в движении частиц: вокруг живых, метаболически активных колоний частицы демонстрировали ускоренное движение, соответствующее направленному перемещению, тогда как вокруг погибших или метаболически неактивных клеток частицы двигались случайно, как при диффузии. Кроме того, когда концентрация глюкозы в среде была снижена ниже порогового уровня, потоки исчезали, демонстрируя прямую связь между метаболизмом и генерацией течений. Это указывало, что величина метаболической активности и размер колонии должны достигать критических значений для возникновения такого эффекта. Как оказалось, существует пороговое значение размера колонии, при котором массовое метаболическое потребление формирует достаточно мощные плотностные градиенты, чтобы индуцировать стабильные и устойчивые течения.
Колонии меньшего размера таких потоков не создавали — движение частиц там соответствовало обычной тепловой диффузии. Более того, если большой кластер был искусственно раздроблен на мелкие фрагменты, каждый отдельный фрагмент терял способность генерировать потоки. Однако когда мелкие кластеры аггрегировали в более крупный, поток появлялся вновь. Это демонстрирует критическую роль именно крупного размера и суммарного метаболического вклада. Дальнейшие эксперименты выявили, что соседствующие крупные кластеры не создают единого объединённого потока, а выступают как независимые «насосы» плотности.
Между ними образуется застойная точка, свидетельствующая о том, что каждый кластер формирует свою локальную среду и локальный поток, привлекая жидкость именно к себе. Такой эффект может иметь значение для понимания взаимодействия микроэкологических систем, когда колонии конкурируют за ресурсы и пространство. Открытие метаболически вызванных потоков у снежных дрожжей принципиально меняет понимание биофизических ограничений эволюции многоклеточности. Ранее считалось, что отсутствие специализированных транспортных систем ограничивает размер колоний исключительно сферическими эффектами и снижает скорость роста — рост становится только линейным или субэкспоненциальным. Реальные наблюдения показывают, что через возникновение простых физических процессов — конвективных потоков — эти ограничения частично устраняются, позволяя экспоненциальный рост и достижение больших размеров без необходимости эволюции сложных биологических транспортеров.
Это свидетельствует, что физические свойства и эффект биофизических факторов, таких как плотностные градиенты и силы тяжести, могут служить так называемым «биофизическим каркасом», облегчающим появление новых биологических функций и направляя эволюцию к более сложным формам жизни. Важно отметить, что структура снежных дрожжей, сформированная в ходе эволюции, представляет собой не просто слипшиеся клетки, а механически стабильные, пористые и переплетённые структуры, которые позволяют жидкости свободно проникать внутрь. Такая архитектура дополнительно способствует возникновению и поддержанию потоков. Если бы колонии были плотными и непроницаемыми, создание активных циркуляций стало бы невозможным. Тем самым сочетание биологической эволюции морфологии и базовых физических принципов обеспечивает непрерывное увеличение размера и сложность колоний.
Это открытие вносит важный вклад не только в микробиологию и биофизику, но и в общую теорию эволюции. Оно подчеркивает, насколько тесно связаны физика и биология: физические законы не только ограничивают, но и открывают новые пути эволюционного развития. Такие биофизические механизмы могут готовить основу для более специализированных адаптаций в будущем, обеспечивая промежуточные этапы, когда новые функции формируются не за счёт генетических изменений, а благодаря физическим свойствам и их взаимодействию с биологией. Перспективы развития исследований в данной области огромны. Вопросы о том, как эти потоки влияют на дифференциацию клеток, на формирование тканей, на обмен сигналами между клетками в колониях, а также возможность генетической ассимиляции этого феномена, остаются открытыми.









